
Latest News
-

Laura and Janaína attend ESA at Salt Lake City, Utah
The Annual Meeting of the Ecological Society of America was held in Salt Lake City, Utah. This year’s theme was Ecology in the Age of Uncertainty Laura presented in the session How Will AI Change Ecology? She talked about some pretty ‘hot-takes’ in the field including: ‘Do we even need to teach students to code?’…
-

Laura attends the WBF
Laura presented at the World Biodiversity Forum in June at Davos, Switzerland to talk about how we can reimagining adaptive sampling.
-

Laura and Isaac attend DeepSDM Day
Laura and Isaac attend DeepSDM Day 2026 at Sion, Switzerland On June 12th. The day featured keynotes (one from Laura!), short talks, and posters that touch on species distribution modelling with deep learning.
-

Welcome Maho!
We’re excited to welcome Maho Horikawa, our new Master’s student joining us this fall. She is one of our stars in the Blitz the Gap project and can’t wait to see what she’ll do next.
-

Blitz the Gap
Another amazing year of blitzing the gap Join us again this summer for the Canadian wide bioblitz
-

Congratulations Dr. Serrano!
Huge congratulations to Dr. Serrano for passing her thesis PhD defence! Her work on global threats to biodiversity is a great addition to the field. We’re so proud of you!
-
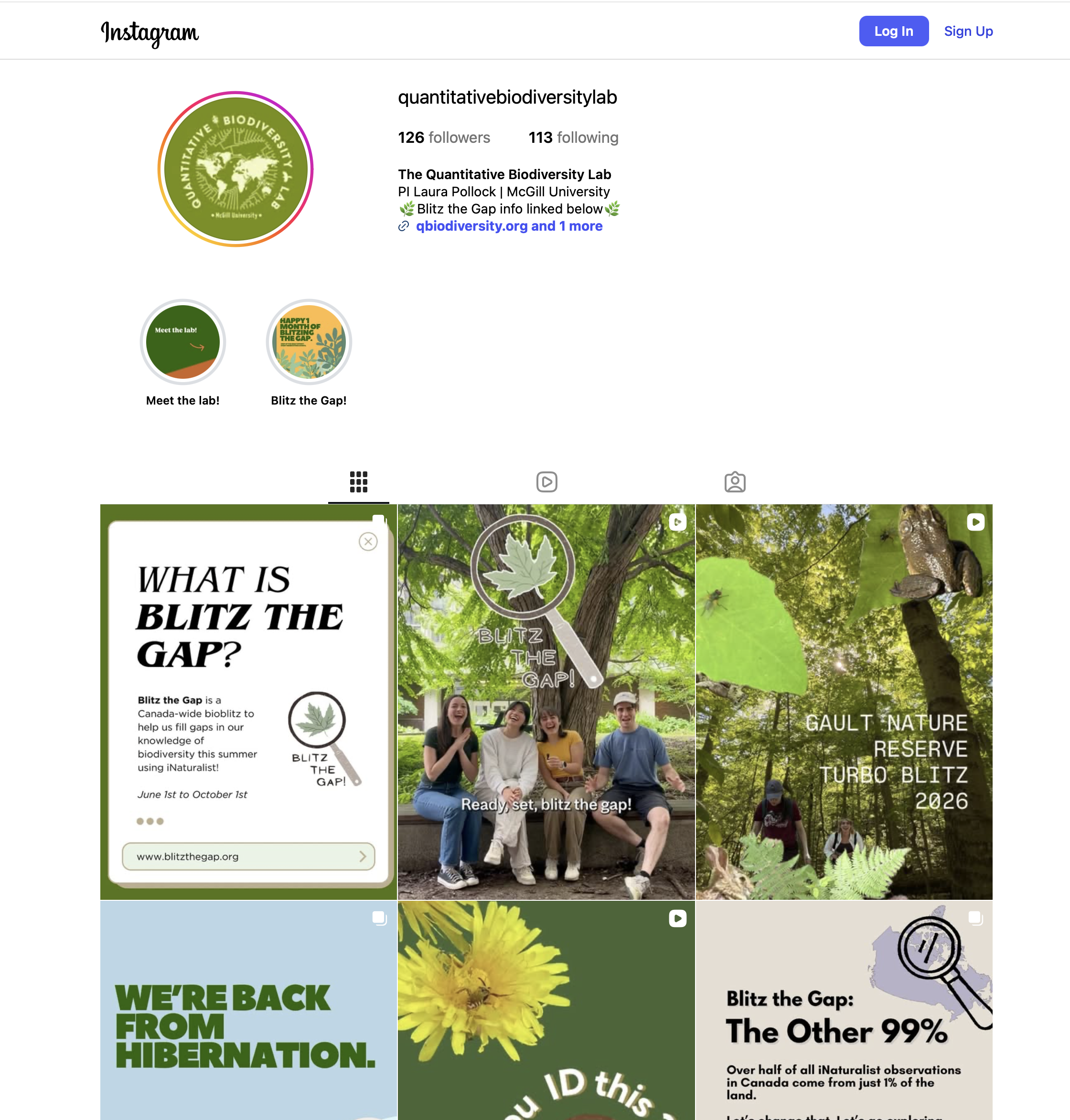
Follow the lab on instagram
We’re excited to launch our new Instagram account @quantitativebiodiversitylab! Follow along for behind-the-scenes looks at blitz the gap, new publications, lab life, and the latest in biodiversity research. Give us a follow and join the community!
-

-

Protecting biodiversity and ecosystem services
The many values of nature: Intrinsic value reflects the high value attributed to biodiversity (e.g. species) and ecosystem functioning, independently of human presence. Cultural value reflects the high value placed on natural heritage and traditions rooted in nature (e.g. the diversity of mushroom and plant species for gathering, heritage landscapes, nature tourism). Regulating value places…
-

adieu la france, bonjour le canada
So, I’m incredibly excited (and a little heartbroken!) to be finally leaving France and completing my full circle around the globe to end up back in North America. I’ll be starting as Asst. Professor in the Biology department at McGill University in Montreal this summer. It’s been an amazing journey and my life has been so…